Большинство позвоночных обладают тремя основными типами проприоцепторов: мышечными веретенами, которые встроены в скелетные мышечные волокна, сухожильными органами Гольджи, которые лежат на границе мышц и сухожилий, и суставными рецепторами, которые являются низкопороговыми механорецепторами, встроенными в суставные капсулы.
Где расположены проприорецепторы?
Проприорецепторы, проприоцепторы, чувствительные рецепторы(от лат. proprius — «собственный, особенный» и receptor — «принимающий») — периферические элементы сенсорных органов, расположенныев мышцах, связках, суставных сумках, в коже и свидетельствующие об их работе (сокращения мышц, изменения положения тела в пространстве).
Что отражает проприорецептивное ощущение?
Среди рецепторов выделяют проприоцепторы, или глубокие рецепторы, которые локализуются в мышцах, сухожилиях, связках и суставах. Они воспринимают такие раздражения, как чувствовеса, давления, положение частей тела в пространстве, степень напряжения мышц [1,2,5]. Таким образом, проприоцепция — это ощущениеположения частей тела и их движения у животных и человека, иными словами — ощущениесвоего тела в пространстве.
Проприорецепция (другое ее название «мышечно-суставная рецепция») – это восприятие проприорецепторами мышц, сухожилий, фасций и суставных сумок, состояния мышц, положения тела в пространстве, передача информации в ЦНС и восприятие ее сенсорными нейронами коры больших полушарий. Проприорецепторы находятся в мышцах, связках, сухожилиях, суставах, глубоких слоях кожи. Они дают представление вместе со зрительным анализатором и вестибулярным аппаратом о положении тела и его частей в пространстве. Это мышечные веретена, сухожильные тельца Гольджи, кожные сухожильные связочные тельца Фатера-Пачини. Все они являются первично чувствующими, медленно адаптирующими механорецепторами. Мышечные веретена обладают спонтанной активностью, что способствует поддержанию тонуса мышц.
Адекватным раздражением проприорецепторов является сокращение скелетных мышц.
Мышечные веретена рассеяны по всем скелетным мышцам. Концы их обычно прикрепляются к мышечным волокнам параллельно. Каждое веретено покрыто капсулой, которая расширятся в центре и образует ядерную сумку. Внутри веретена содержатся интрафузальные мышечные волокна. Эти волокна в 2-3 раза тоньше обычных (экстрафузальных) волокон скелетных мышц, на них спирально расположены окончания афферентных волокон
Интрафузальные волокна подразделяются на два типа:
1) Длинные и толстые (диаметр 20-25 мкм), которые информируют ЦНС о динамическом компоненте движения – скорости изменения длины мышцы. Таких волокон в мышечном веретене не более двух.
2) Короткие и тонкие (диаметр 10–12 мкм), которые информируют ЦНС о статическом компоненте движения – текущей длине мышцы. Таких волокон в мышечном веретене от 2 до 12.
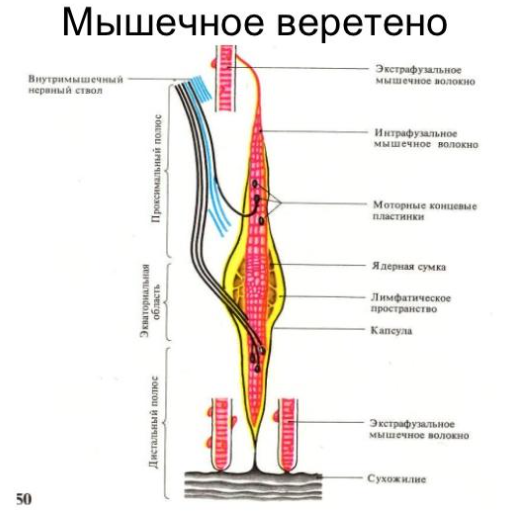
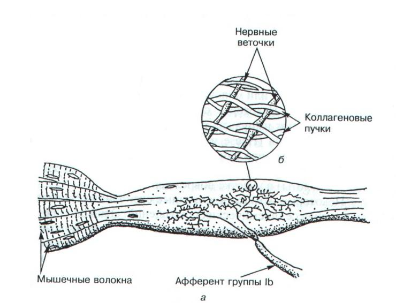
Веретено имеет и эфферентную иннервацию, к нему идут аксоны гамманейронов, которые расположены в спинном мозге. Проксимальный конец веретена прикрепляется к волокну скелетной мышцы, а дистальный – к фасции. Веретено растягивается при расслаблении скелетной мышцы, происходит генерация импульсов, которые по афферентным волокнам поступают в спинной мозг и возбуждают мотонейроны своей мышцы, а через тормозящий нейрон вызывают торможение мотонейронов мышцы-антагониста. Возбуждение альфа-мотонейронов, которые иннервируют скелетные мышцы, сопровождается возбуждением гамма-мотонейронов, иннервирующих веретена. Сухожильные рецепторы Гольджи: Рецепторы Гольджи располагаются в месте перехода мышечных волокон в сухожилия. Когда мышечные волокна сокращаются, коллагеновые волокна натягиваются и сжимают нервные веточки, которые начинают импульсировать (рис.4). Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. Сухожильные рецепторы возбуждаются в 1,5 – 8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении.
Суставные рецепторы: Суставные рецепторы подразделяются на несколько типов в зависимости от их реакции на амплитуду, скорость и направление движения в суставе.
1) Тельца Руффини находятся в капсуле сустава и воспринимают направление и скорость изменения межзвенного угла. Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
2) Тельца Пачини посылают в ЦНС информацию о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы посылают в ЦНС информацию о значениях межзвенных углов, то есть о положении сустава. Их импульсация продолжается в течение всего периода сохранения межзвенного угла, и она тем больше, чем больше изменения угла.
Вестибулорецепция
Вестибулорецепция обеспечивает восприятие положения головы и всего тела в пространстве, прямолинейных или вращательных ускорений при движении тела (акселерационное чувство), участвует в регуляции тонуса мышц для поддержания естественной позы и восстановления нарушенной позы, а также принимает участие в координации движений. Вестибулорецепция обеспечивается вестибулярной сенсорной системой и вестибулорецепторами. Это рецепторы трех полукружных каналов лабиринта пирамиды височной кости и отолитового аппарата в преддверии улитки.

Вестибулорецепция обеспечивается вестибулярной сенсорной системой. Периферическим отделом ее является вестибулярный аппарат — три полукружных канала и отолитовый аппарат.
Полукружные каналы располагаются в трех взаимно перпендикулярных плоскостях. Один из концов каждого канала расширен (ампула). Полукружные каналы повторяют форму костных каналов; они перепончатые, заполнены плотной эндолимфой, только в ампулах располагаются рецепторные волосковые клетки.
Отолитовый аппарат находится в двух мешочках преддверия улитки. Первый мешочек лежит ближе к улитке, а второй — к полукружным каналам. Отолитовый аппарат — скопление рецепторных волосковых клеток. В полость мешочка выступает часть рецепторных клеток с одним более длинным волоском и 60… 80 склеенными неподвижными волосками. Волоски пронизывают желеобразную мембрану, содержащую кристаллики карбоната кальция – отолиты.
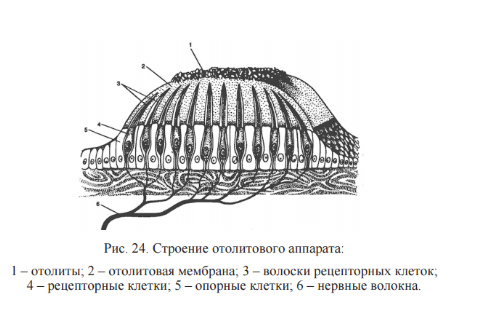
Изменение положения головы, ускорение и замедление движения, тряска, качка вызывают движение эндолимфы в полукружных каналах и скольжение отолитовой мембраны по волоскам, что сопровождается сгибанием волосков и генерацией в них импульсов. Импульсы через синапсы посредством ацетилхолина передаются на окончания волокон вестибулярного нерва, во многие отделы центральной нервной системы: в кору больших полушарий, ретикулярную формацию, мозжечок, глазодвигательные ядра, средний мозг, продолговатый мозг. Таким образом обеспечивается динамическое перераспределение тонуса скелетных мышц.
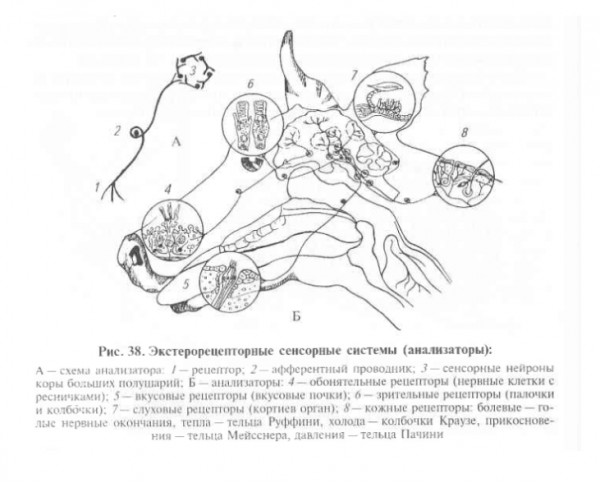
Экстерорецепторы и экстерорецепция
Экстерорецепторы (от лат. exter – внешний) – группа специализированных чувствительных образований, воспринимающих раздражения, воздействующие на организм из окружающей его внешней среды (антагонисты – интерорецепторы).
Экстерорецепцию подразделяют на 8 видов: болевая, температурная, прикосновения, давления, вкусовая, обонятельная, слуховая и зрительная (в качестве иллюстрации – рисунок в начале страницы).
Болевая рецепция
Болевая (ноцицетивная) система представляет собой совокупность нервных образований, обеспечивающих формирование болевых ощущений и их угнетение.
Боль – это неприятное, в виде страдания, ощущение, возникающее в результате действия на организм сверхсильного раздражителя, патологического процесса или кислородного голодания. Болевой раздражитель несет мало информации об окружающем мире, он играет защитную роль, предупреждая организм о действии вредного фактора.
Восприятие боли:
Периферический отдел болевой системы представлен рецепторами боли – ноцицепторами (от лат. nocere – вредить, разрушать), которые реагируют на стимулы, угрожающие организму повреждением или вызывающие повреждения. Они найдены в коже, слизистых, серозных оболочках, надкостнице, зубах, мышцах, органах грудной и брюшной полости. Болевые рецепторы всех органов и тканей представляют неинкапсулированные окончания нервных волокон, имеющие форму волосков, пластинок, спиралей. На поверхности кожи число ноцицепторов (болевых точек) намного превышает число тактильных и температурных рецепторов.
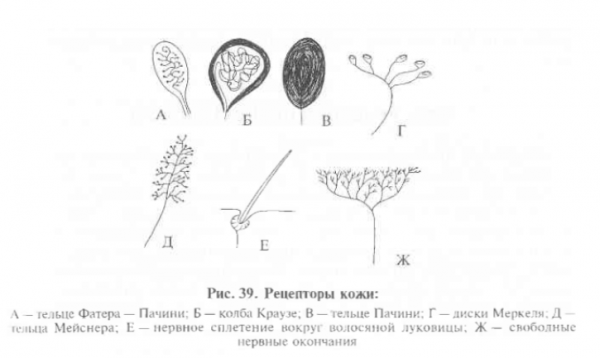
Различают 4 типа ноцицепторов:
• Механоцицепторы – они возбуждаются очень сильными ме-
ханическими раздражителями, вызывающими деформацию
и повреждение мембраны рецептора при сжатии или растя-
жении ткани;
• Хемоноцицепторы – для них специфическими раздражи-
телями являются алгогены – вещества, выделяющиеся
при повреждении клеток или развитии воспалительного
процесса в тканях. Различают 3 типа алгогенов: тканевые
(ацетилхолин, серотонин, гистамин, ионы K+, Na+, H+),
плазменные (брадикинин, каллидин, лейкотриины) и вы-
деляющиеся из нервных окончаний (вещество П, ВИП –
вазоинтестинальный пептид). Алгогены различных групп
обладают различными механизмами активации болевых
рецепторов: тканевые активируют непосредственно боле-
вые окончания, плазменные – также могут прямо воздействовать
на нервные окночания, но чаще они повышают
чувстивтельность ноцицепторов. Вещество П (медиатор
боли) при повреждающих воздействиях выделяется из не-
рвных окончаний и действует на рецепторы, локализован-
ные на этих окончаниях, вызывая генерацию потока ноци-
цептивных импульсов.
• Термоноцицепторы – воспринимают боль при изменении
температуры выше 43 градусов или ниже 17 градусов
• Полисенсорные – механо-, термо- и хемоноцицепторы. Они
встречаются наиболее часто.
Температурная рецепция
Терморецепторы — это вид рецепторов, который воспринимает температурные изменения.
Терморецепторы:
• периферические – расположены в коже, слизистых оболоч-
ках, роговице глаза;
• висцеральные – во внутренних органах, сосудах, много в ды-
хательных путях;
• центральные – в основном в гипоталамусе (это нейрорецеп-
тивные нейроны).
Специфические терморецепторы кожи представлены холодо-
выми (колбочки Краузе) и тепловыми (тельца Руфини) нервными
окончаниями, которые также несут механорецепторные функции. Рецепторы холода лежат поверхностно (0,17 мм), а рецепторы тепла – глубоко (0,3 мм) в коже.
Формы температурных ощущений – холод, тепло.
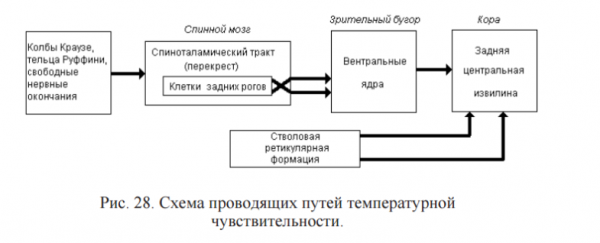
Рецепция прикосновения
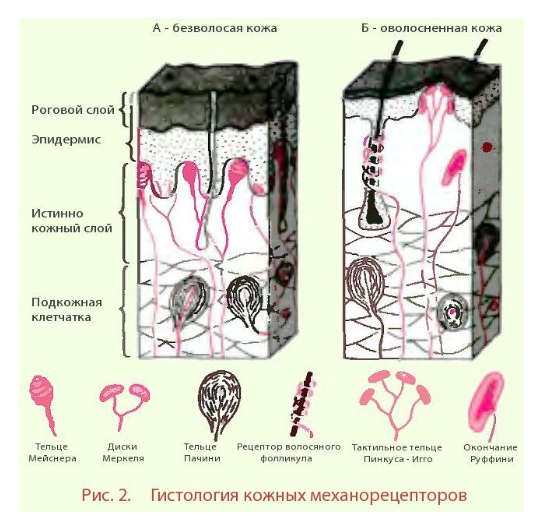
Рецепция прикосновения (тактильная рецепция) – это восприятие рецепторами прикосновения (мейснеровы тельца и меркелевы диски) предметов, передача информации в ЦНС и восприятие ее сенсорными нейронами коры. Рецепторы прикосновения расположены в коже (рис. ниже), в слизистой оболочке языка.
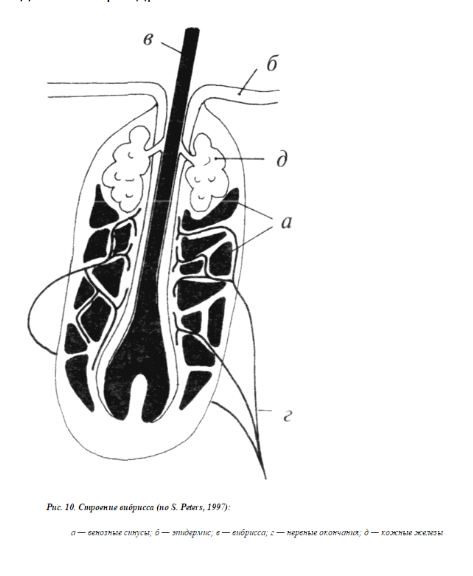
У животных есть еще специальные осязательные волоски – вибриссы, расположенные на морде. Эта рецепция обеспечивает приспособительные реакции в виде смещения кожи, различных движений (перемещений) в целях устранения или избегания действия раздражителя.
Рецепция давления
Рецепция давления – восприятие рецепторами давления (тельца Пачини) раздражителей, которые вызывают деформацию кожи, передачу информации в ЦНС и восприятие ее сенсорными нейронами коры. Они находятся в глубине кожи губ, носа, спины, живота, шеи и других участках тела. Отличительное свойство рецепторов давления – свойство более высокой адаптации. Эта рецепция обеспечивает защитные двигательные реакции.
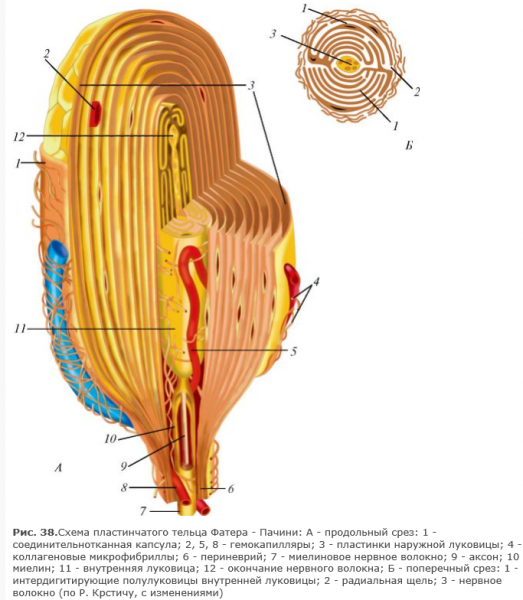
Вкусовая рецепция (анализатор)
Вкус, так же, как и обоняние, основан на хеморецепции. Вкусовые рецепторы несут информацию о характере и концентрации веществ, поступающих в рот. Их возбуждение запускает сложную цепь реакций разных отделов мозга, приводящих к различной работе органов пищеварения или удалению вредных для организма веществ, попавших в рот с пищей.
Рецепторы вкуса — вкусовые почки — расположены на языке, задней стенке глотки, мягком небе, миндалинах и надгортаннике. Их количество исчисляется миллионами (например, у собак – свыше 200 млн.). Больше всего их на кончике языка, его краях и задней части. Каждая из вкусовых почек состоит из нескольких рецепторных клеток и, кроме того, из опорных клеток. Вкусовая почка имеет колбовидную форму, она не достигает поверхности слизистой оболочки языка и соединена с полостью рта через вкусовую пору, поэтому только после растворения вещество проникает через поры и воспринимается рецепторами. Каждая из рецепторных вкусовых клеток имеет на конце 30-40 микроворсинок, которые воспринимают химические вещества. Вкусовых раздражителей – 4 основных вида: сладкое, кислое, соленое и горькое. Кончик языка наиболее чувствителен к сладкому, боковые поверхности – к соленому и кислому, корень – к горькому.
Вкусовая рецепция обеспечивает вкусовые ощущения и приспособительные пищевые реакции, как: образование и выделение слюны, желудочного и других пищеварительных соков, сокращение желудка и кишечника
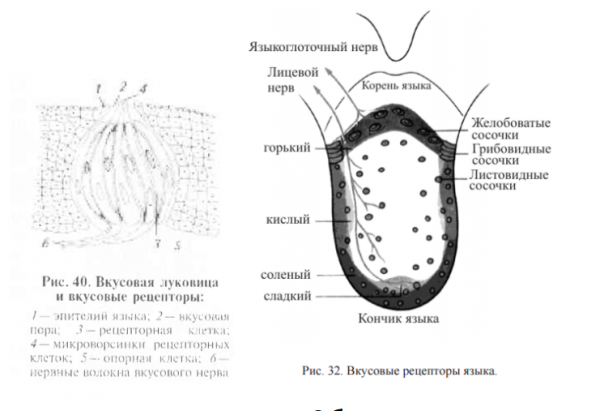
Обонятельная рецепция
Обонятельные рецепторы воспринимают химические раздражители из внешней среды. Система обоняния способствует ориентации в окружающей среде и познании ее, участвует в формировании
пищевого поведения (выработка условных пищевых рефлексов, избегание и избавление от неприятных раздражителей), в формировании эмоций.
На запахи реагируют обонятельный эпителий верхней задней полости носа с обонятельными рецепторами (это первично-чувствующие хеморецепторы), а также свободные окончания тройничного нерва.
Обонятельный рецептор представляет собой булавовидное окончание короткого дендрита обонятельной клетки, содержащее 6–12 ресничек с рецепторными белками, взаимодействие которых с пахучими веществами обеспечивает формирование обонятельных ощущений.
Известно более 100 первичных обонятельных ощущений, которые объединяются в 7 групп основных различаемых запахов – камфорный,
цветочный, мускусный, мятный, эфирный, едкий, гнилостный. В естественных условиях воспринимаются смешанные запахи. Каждый
обонятельный рецептор возбуждается при действии многих пахучих
веществ, но особенно тех, к которым он наиболее чувствителен.
Проводниковый отдел обонятельного анализатора начинается
аксонами биполярных обонятельных нейронов, которые пронизывают пластинку решетчатой кости и формируют обонятельный нерв. Волокна обонятельного нерва образуют синапсы с крупными
митральными клетками обонятельной луковицы. Аксоны митральных клеток формируют обонятельный тракт. Волокна обонятельного тракта отдельными пучками идут в передние ядра таламуса,
а затем в кору (некоторые авторы считают, что отростки второго
нейрона идут прямо в кору большого мозга, минуя таламус).
Корковый отдел обонятельного анализатора представлен обонятельной извилиной, гиппокампом, прегрушевидной областью. Нейроны, отвечающие на обонятельные стимулы обнаружены также
в ретикулярной формации, имеется связь с вегетативными ядрами
гипоталамуса. Тесная связь с лимбической системой объясняет присутствие эмоционального компонента в обонятельном восприятии.
Адаптация к действию пахучего вещества в обонятельной системе происходит в течение десятка секунд или минут, зависит от
скорости потока воздуха и концентрации пахучего вещества. При-
чем сами обонятельные нейроны продолжают генерировать ПД. Это
указывает на то, что развитие адаптационных процессов происходит
в проводниковом и корковом отделах обонятельного анализатора.
Особое сигнальное значение имеют пахучие вещества, выделяемые животными – телергены (действующие вдали). Гомотелергены – феромоны. Они являются средством общения особей одного вида, сигнализируют о поле, возрасте, функциональном состоянии животного. Животных с хорошо развитым обонянием называют макросматиками (большинство млекопитающих, некоторые придонные рыбы, акулы и др.). У макросматиков реснички обонятельных рецепторов высоко подняты над поверхностью обонятельного эпителия, значительное число обонятельных клеток и ресничек на них увеличивает площадь обонятельной поверхности.
Микросматики – животные со слабой степенью развитости обонятельной рецепции (птицы, приматы). Это связано со снижением числа обонятельных рецепторов и ресничек. Аносматики – киты, дельфины – не имеют обонятельных рецепторов и луковиц. Обонятельный анализатор обеспечивает целый ряд приспособительных поведенческих реакций: пищевых, половых, оборонительных и исследовательских.
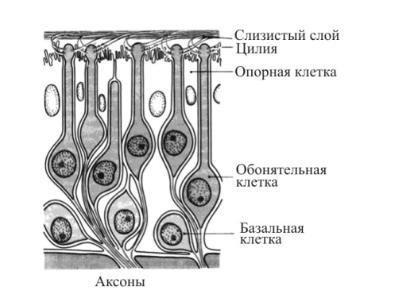
Слуховая рецепция
Слуховой анализатор – второй по значимости после зрительно-
го. С помощью слуха живые организмы ориентируются в звуковых
сигналах окружающей среды, формируют соответствующие пове-
денческо-приспособительные реакции: например, пищедобыва-
тельные и оборонительные. У человека слух играет важную роль
в связи со способностью восприятия разговорной и вокальной речи,
музыкальных произведений, является необходимым компонентом
общения и приспособления.
Периферический отдел слухового анализатора представлен пар-
ным образованием – органом слуха, который состоит из наружного,
среднего и внутреннего уха.
Адекватным раздражителем для слуховой системы являются
звуки, т.е. колебательные движения частиц упругих тел, распространяющихся в виде волны в самых различных средах, включая
воздушную.
Наружное ухо – раковина, наружный слуховой проход –
улавливает направление звука (биуральный слух – слух двумя
ушами), концентрирует, усиливает, проводит как рупор звуковые
колебания к барабанной перепонке, а также защищает её от механических и температурных воздействий внешней среды. Барабанная перепонка отделяет наружное ухо от среднего, по форме напоминает пологую воронку, служит для точной передачи звуковых колебаний в среднее ухо.
Среднее ухо – барабанная полость с воздухом, в которой последовательно расположены 3 слуховые косточки: молоточек вплетен своей рукояткой в барабанную перепонку, а другим концом связан через суставную поверхность с наковальней, которая сочленена со стремечком, прилегающим к мембране овального окна. Слуховые косточки проводят звуковые колебания к внутреннему уху, значительно усиливая их. Это возможно благодаря большей площади барабанной перепонки по сравнению с овальным окном и за счет того, что барабанная перепонка передает свои колебания на более длинное плечо рычага, образованного слуховыми косточками.
Интенсивность звукового давления меняется с помощью защитного рефлекторного механизма, который представлен сокращением двух мышц – мышцы, натягивающей барабанную перепонку, и мышцы, фиксирующей стремечко. Сильные звуки активируют напряжение этих мышц, что ограничивает амплитуду колебаний барабанной перепонки и движение стремечка, предохраняя тем самым рецепторный аппарат внутреннего уха от чрезмерного возбуждения и разрушения. Этот механизм не успевает срабатывать в ответ на мгновенные сильные раздражители – взрыв, выстрел, сильный звук колокола.
Среднее ухо соединяется с задней частью глотки евстахиевой трубой, которая служит для выравнивания давлений в барабан-
ной полости и наружном слуховом проходе, а также выполняет
вентиляционную и дренажную функции. В состоянии бодрство-
вания евстахиева труба открывается каждую минуту, а также при
жевании, глотании, зевании; а во время сна – каждые 5 минут. Если внешнее давление изменяется быстрее (во время наращивания вы-
соты самолетом или при его снижении), а глотание не происходит,
то разность давлений в барабанной полости и в окружающей среде
приводит к снижению восприятию звуков вследствие натяжения
барабанной перепонки («закладывание ушей»).
Звуки проводятся также по костям черепа. Исследование воз-
душной и костной проводимости у пациента помогает врачу в пос-
тановке диагноза. Таким образом, наружнее и среднее ухо – это
звукоулавливающий и звукопроводящий аппараты слуховой системы.
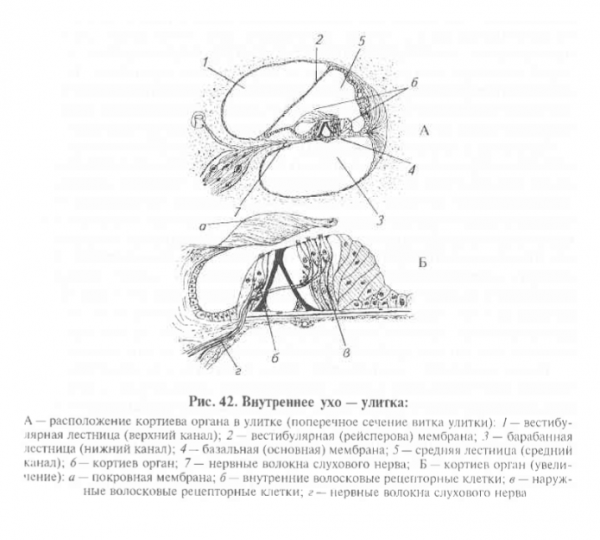
Внутреннее ухо расположено в пирамидке височной кости. Там
находится звуковоспринимающий рецепторный аппарат: улитка
с кортиевым органом. Вестибулярная и базальная мембраны разде-
ляют улитку на 3 канала – вестибулярную, барабанную и среднюю лестницы. Вестибулярная и барабанная лестницы заполнены перилимфой (она напоминает спинно-мозговую жидкость). На вершине улитки эти каналы соединяются между собой через отверстие – геликотрему. Средний перепончатый канал изолирован и заполнен эндолимфой (в ней в 100 раз больше К+ и в 10 раз меньше Nа+ , чем в перилимфе, выше вязкость). В среднем канале на базальной мембране расположен кортиев орган с рецепторами. Фонорецепторы – это вторично чувствующие механорецепторы, представленные волосковыми клетками. Кортиев туннель разделяет волосковые клетки на две группы – внутреннюю и наружную. Внутренние клетки образуют один ряд, а наружные – 3–5 рядов.
Волосковые клетки покрыты сверху текториальной (покровной)
мембраной, которая сгибает волоски (цилии), погруженные в ее
вещество. Вестибулярная лестница сообщается со средним ухом
через овальное окно, к его мембране прилежит стремечко. Барабанная лестница сообщается со средним ухом через круглое окно, закрытое мембраной. Наличие овального и круглого окон обеспечивает возможность колебания перилимфы. При этом, когда толчок стремечка передается на мембрану овального окна, возникает бегущая волна перилимфы, которая достигает круглого окна, смещая его мембрану в полость среднего уха.
Зрительный анализатор
Зрительный анализатор представляет совокупность структур, воспринимающих световые волны и формирующих зрительные ощущения.
Рецепторный аппарат глаза расположен в сетчатке. Сетчатка представляет собой сложную многослойную структуру, объединяющую рецепторы и нейроны.
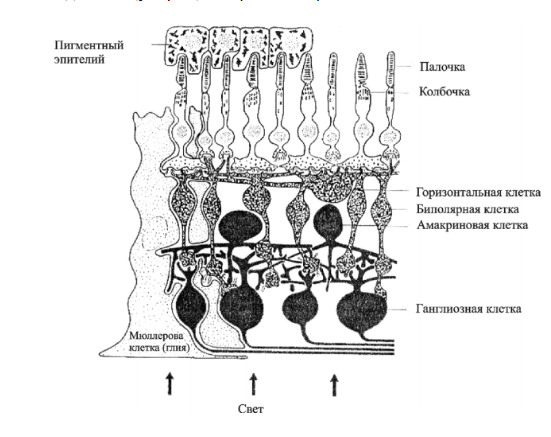
Фоторецепторы сетчатки – палочки и колбочки, локализованы в ее пигментном слое, наиболее удаленном от хрусталика. Этот слой образован эпителием, содержащим фусцин. Последний поглощает свет, препятствуя его отражению и рассеиванию, что обеспечивает четкость зрительного восприятия.
Отростки пигментных клеток окружают светочувствительные членики палочек и колбочек. Они принимают участие в обмене веществ фоторецепторов и в синтезе зрительных пигментов, фагоцитируют продукты распада зрительных пигментов.
Палочек больше на периферии, их функция – восприятие света, периферическое, сумеречное зрение. Колбочки находятся в центре (желтое пятно с центральной ямкой), их функции – восприятие цвета, центральное, дневное зрение. Белое пятно на сетчатке соответствует месту выхода зрительного нерва, там нет фоторецепторов.
Зрительная рецепция связана с деятельностью вспомогательных приспособлений, которые вместе со зрительными рецепторами формируют глаз. Вспомогательные системы глаза – это оптическая система и защитный аппарат. 1) Оптическая система глаза представлена передней и задней поверхностью роговой оболочки, хрусталиком и стекловидным телом. Для ясного видения предмета необходимо, чтобы лучи от всех его точек падали на сетчатку. Приспособление глаза к ясному видению разноудаленных предметов называют аккомодацией. Аккомодация осуществляется путем изменения кривизны хрусталика. Рефракция – преломление света в оптических средах глаза.
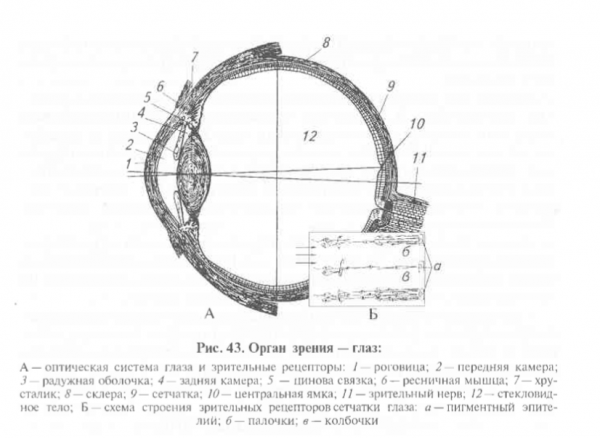
Свет, воспринимаемый животными, представляет электромагнитные излучения в диапазоне 400-700 нм. Основа зрения – восприятие контраста между светлым и темным. Количество света, падающего в глаз, регулируется зрачком (отверстие в радужной оболочке). Контроль за изменением размера зрачка осуществляется автоматически нервными волокнами, заканчивающимися в мышцах радужной оболочки. Суживающие зрачок циркулярные мышцы иннервируются
парасимпатическими волокнами, а расширяющие его радиальные
мышцы – симпатическими. Зрачок ведет себя как диафрагма – регулирует поток света (на свету суживается, в темноте расширяется), уменьшает сферическую абберацию глаза, пропуская те лучи, которые попадают на центральную часть хрусталика, где фокусировка более точная. 2) Защитный аппарат глаза включает в себя верхние и нижние веки, 3-е веко, слезный аппарат, который обеспечивает защиту фоторецепторов от чрезмерного света, мейбомиевы железы, выделяющие глазную смазку. Со слезной рецепцией связаны следующие приспособительные реакции: пищевые, комфортные, оборонительных, исследовательских, коммуникационных и др.

